

麦角硫因是当下市场最热门的天然活性成分之一。作为一种独特的天然抗氧化剂和细胞保护剂,麦角硫因在化妆品领域中具有多种功效,如抗氧化、抗衰老、抗炎、抗糖、美白、防晒等。麦角硫因可以通过OCTN-1转运蛋白特异性运输和积累,特别是在线粒体和细胞核中,因此其能在人体全身组织内积累并发挥作用,在医药、食品、化妆品等多领域都有广泛用途。欧美和日本均认可麦角硫因为安全食品原料,极具应用前景。本文对麦角硫因在化妆品方面的功效研究和实际应用做了梳理和分析,为麦角硫因在化妆品领域的研发和应用提供了数据支持和理论支撑。
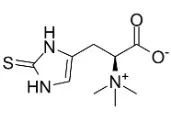
麦角硫因结构式
麦角硫因是机体内的重要活性物质,但人类、动物及高等植物体自身无法合成麦角硫因,必须通过饮食等方面从外界环境中摄取[1]。
麦角硫因最早是Charles Tanret在1909年从感染麦角菌(Claviceps purpurea)和其他真菌的黑麦粒中分离得到。国内外研究表明它在生物体内可起到很强的抗氧化作用[2,3],具有抗炎[4]、抗光老化[5]、螯合二价金属离子、激活抗氧化物酶、维持DNA生物合成、提高细胞免疫[6]等多种生理功能,广泛应用于医药、保健品、食品、饮料、动物饲料、化妆品及生物化工等领域。它作为一种人体内具有专属转运蛋白OCTN-1的天然抗氧化剂,同时具有高效、安全、无毒的特性,已经走入人们的视野,成为研究的热点。
麦角硫因由于具有显著的生物学功能,各国学者很早就对其展开了研究。
麦角硫因的制备方法有三种:化学合成法、提取法[7]和生物发酵合成法[8]。法国OXIS公司第一个研制出了高效合成麦角硫因的方法,并于1995年申请了专利,通过巯基导入咪唑环制备;但化学合成法的安全性难以得到保证,且合成原料昂贵、售价居高。提取法是从许多大型真菌的子实体中提取麦角硫因,原料含量低、杂质多、提取成本也很高。生物发酵法则利用微生物代谢转化生成,近年来越来越多通过基因重组和代谢改造来获取麦角硫因的研究得到了实质性的进展,纯度和生产效率大幅提升[9-11],国内以中科院上海有机所发明的麦角硫因的制备方法为典型。
本文对麦角硫因的全球专利申请进行统计和分析,对其在化妆品领域的研究和应用进行梳理和总结,以期为我国麦角硫因产业和实际应用做好铺垫,同时为中国高端的功效型化妆品的崛起做出一份贡献。
1.麦角硫因全球专利申请概况
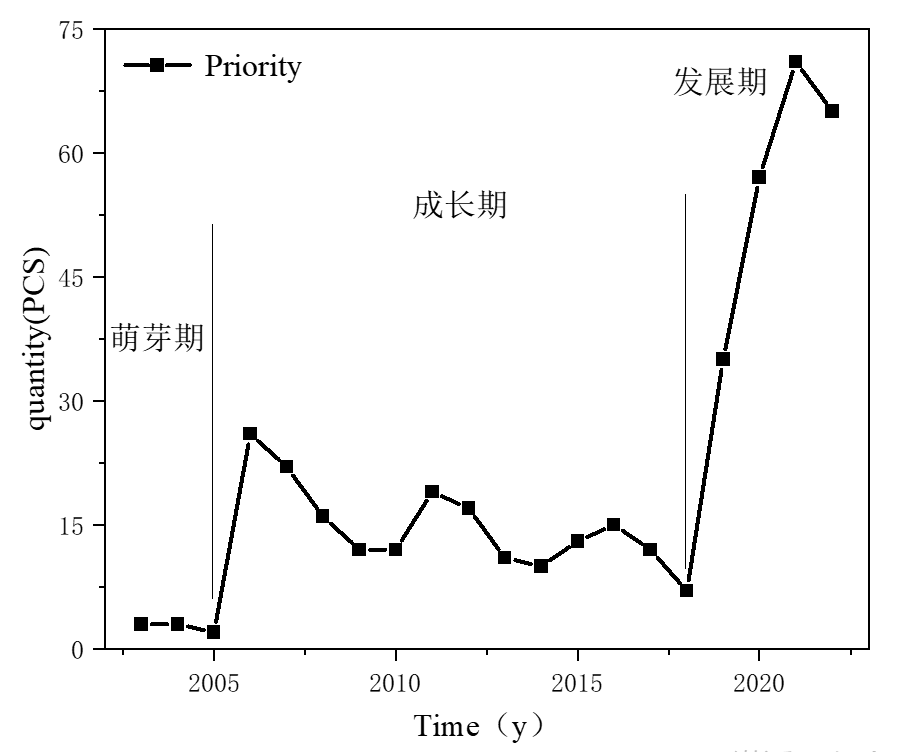
麦角硫因专利申请量与年份关系图
LVMH研究所认为在麦角硫因和抗坏血酸2-葡糖苷比值在0.005~60时,再添加美容领域常用的佐剂制备成乳液、凝胶、精华素、贴剂或面膜,将这些化妆品涂抹于皮肤可刺激修复真皮和表皮细胞DNA中氧化碱基的酶(CN200810182711.X)。
2.2 麦角硫因对线粒体的修复作用
近年来,线粒体已被认定是许多药物毒性的重要靶标,细胞的生死存亡很大程度上取决于线粒体的功能状态。线粒体有氧代谢电子传递链的副产物(ROS)的积累可引起细胞氧化应激,从而导致蛋白质、脂质和细胞器的损伤。线粒体损伤会产生更多的ROS不能及时被消除、产生积聚、形成恶性循环,持续损伤线粒体直至细胞功能缺失甚至凋亡等。
麦角硫因可保护线粒体免受自然或疾病过程以及外源因素的影响(US6479533B1)。使用磷脂酰胆碱、磷脂酰乙醇胺、油酸和半琥珀酸胆固醇酯组成的脂质体包裹麦角硫因处理细胞发现该脂质体可以保护线粒体免受自然和疾病过程中造成的氧化损伤,并减少紫外线和氧自由基造成的损伤。此外,麦角硫因的功效和递送通过封装在脂质体中得到增强。在麦角硫因浓度范围为50 μM至5 mM时,其组合物可以保护线粒体免受辐射、自由基和活性氧等物质的损害;通常以12 μM浓度的麦角硫因脂质体组合物制备成化妆水凝胶局部施用于皮肤表面(US006103746A)。
2.3麦角硫因在细胞中诱导Nrf2活性的用途
皮肤是人体最大的器官,是人体隔离外部损伤的重要屏障。来自太阳的紫外线辐射是环境引起皮肤损伤的主要原因之一,会导致红斑或者晒伤。长期暴露于紫外辐射会导致皮肤光老化、产生自由基、造成急性或慢性损伤,使皮肤晒黑并降解胶原蛋白等。
研究表明麦角硫因能通过诱导细胞中Nrf2的活性,与抗氧化反应元件(ARE)结合,促进下游抗氧化酶的基因表达(TW201549696B,US20160067221A1)。实验表明,使用500 nM麦角硫因孵育细胞1小时可显著提高Nrf2的蛋白总水平,其转录因子调节的HO-1,NQO-1和y-GCLC蛋白的表达也得到增强。因此,麦角硫因在浓度125 nM至500 nM时,可激活人角质形成细胞的Nrf2信号通路,促进Nrf2核易位,增强下游酶促抗氧剂和非酶促抗氧剂的表达,增加细胞的抗光氧化能力,同时抑制UVA刺激造成的细胞凋亡(CN105982890A)。
2.4麦角硫因抑制皮肤蛋白糖化作用
蛋白质的糖化是一种非常普遍的现象,在皮肤老化过程中,胶原蛋白的物理化学性质发生变化,造成皮肤中糖化产物含量增加,最终导致组织僵化并加速皮肤衰老。研究发现(US20020042438A1,JP2001191929A),麦角硫因及其衍生物具有减少甚至抑制蛋白质糖化现象,首先作用于与年龄相关的皮肤张力丧失,其次作用于皮肤的“橙皮”外观。将麦角硫因及其衍生物,以及它们的混合物用于化妆品,如凝胶水溶液、化妆水、乳液等,当麦角硫因含量为0.001-10%时,可以有效减少胶原蛋白的糖化,用于预防和治疗糖化相关的皮肤张力丧失和皮肤老化迹象(EP1166768A1)。
2.5麦角硫因抗光污染、光损伤作用
光污染是现代社会产生的过量或者不适当的光辐射对人类生活和生产环境所造成的的不良影响的现象。紫外线和蓝光都会使人体细胞产生大量活性氧,当体内的氧化程度超出对氧化物的清除速度时,细胞内氧化与抗氧化系统失衡,高水平ROS压迫细胞,可使DNA损伤、脂质过氧化、胶原蛋白结构破坏和线粒体功能损伤,最终导致皮肤出现各种皮肤问题或者皮肤疾病。
已有公开的含有麦角硫因护肤组合物的专利中表示其麦角硫因成分可能具有改善细胞因紫外线、蓝光照射后的细胞缺氧状态,帮助细胞恢复正常的新陈代谢功能,能有效阻拦紫外线、蓝光照射到皮肤上,减少紫外线和蓝光对皮肤的损伤,并促进光损伤后皮肤的修复作用(CN111557875A)。另外有发明表示抗光污染、光损伤组合物中所述的细胞修复组合物包括麦角硫因、泛醇、尿素囊,其中麦角硫因是人体内的重要活性物质,具有清除自由基、解毒、维持DNA的生物合成、细胞的正常生长和细胞免疫等多种生理功能,可以阻挡紫外线侵害皮肤,修复受伤胶原蛋白和弹性纤维,收缩毛孔和防晒等相关作用(CN113855608A)。
2.6 麦角硫因美白、抑制黑色素生成作用
皮肤中的黑色素是提供紫外线保护的天然屏障,在保护皮肤健康方面起着至关重要的作用。但黑色素过多或过少都会引起皮肤颜色异常和皮肤病变,如老年斑和黄褐斑。因此,具有美白祛斑功能的护肤品在化妆品中占有重要地位。麦角硫因抑制黑色素生成或去除黑色素的作用机制可能是由于其独特的结构,即硫取代的咪唑环[13]。含硫醇化合物被认为是重要的黑色素生成抑制剂,因为它们与多巴醌反应形成无色的共轭物。
Liao等人[12]证明了麦角硫因可以与游离酶的底物位点和酶-底物复合物的不同底物位点结合。已有研究证实了麦角硫因对酪氨酸酶的抑制作用优于常见的抑制剂,如曲酸和熊果苷[13-14]。并且存在已经应用到饮品及个人护理物的发明表示,麦角硫因能够抑制人体内酪氨酸酶活性,具有焕颜赋能的效果(CN111388349A,CN104080438B,CN110742822A)。
3. 结 语
麦角硫因,由于其广泛的生理学特性,例如天然、安全、在人体内有专属的转运蛋白、并能直达线粒体内部清除过量自由基和ROS的特点,近几年来快速流行并成为化妆品行业高端、热门的抗衰活动原料之一,受到越来越多的品牌商和生产商的追捧。随着生物合成技术的不断进步及化妆品原料功效测定方法的不断完善,将进一步探索和拓展有关麦角硫因的提取、制备、机理和应用,逐渐缩短与国外的技术和产业差距。本文综述了麦角在化妆品方面的相关专利技术,为后续技术发展和实际应用做好铺垫。
[4] 覃成. BV2细胞模型研究OCTN1转运体功能及其介导抗炎/抗氧化作用机制[D].华中科技大学,2021.DOI:10.27157/d.cnki.ghzku.2021.004888.
[5] Tsay Gregory J.,Lin ShinYi,Li ChienYu,Mau JengLeun,Tsai ShuYao. Comparison of Single and Combined Use of Ergothioneine, Ferulic Acid, and Glutathione as Antioxidants for the Prevention of Ultraviolet B Radiation-Induced Photoaging Damage in Human Skin Fibroblasts[J]. Processes,2021,9(7).
[6] Ko Hyun Ju,Kim Jeongtae,Ahn Meejung,Kim Jin Hwa,Lee Geun Soo,Shin Taekyun. Ergothioneine alleviates senescence of fibroblasts induced by UVB damage of keratinocytes via activation of the Nrf2/HO-1 pathway and HSP70 in keratinocytes[J]. Experimental Cell Research,2021,400(1).
[7] 冯路路,鄂恒超,张艳梅,李晓贝,周昌艳,赵晓燕,任佳丽.食用菌中麦角硫因提取分离和检测方法研究进展[J].食用菌学报,2021,28(01):115-123.DOI:10.16488/j.cnki.1005-9873.2021.01.016.
[8] Qiu Yibin,Chen Zhonglin,Su Erzheng,Wang Libin,Sun Liang,Lei Peng,Xu Hong,Li Sha. Recent Strategies for the Biosynthesis of Ergothioneine.[J]. Journal of agricultural and food chemistry,2021.
[9] Hoek Steven A.van der,Rusnák Matej,Wang Guokun,Stanchev Lyubomir Dimitrov,de Fátima Alves Luana,Jessop Fabre Mathew M.,Paramasivan Kalaivani,Jacobsen Irene Hjorth,Sonnenschein Nikolaus,Martínez José L.,Darbani Behrooz,Kell Douglas B.,Borodina Irina. Engineering precursor supply for the high-level production of ergothioneine in Saccharomyces cerevisiae[J]. Metabolic Engineering,2022,70(prepublish).
[10] 王丽,王阳,李江华,堵国成,康振.产麦角硫因大肠杆菌工程菌株的构建与优化[J].生物工程学报,2022,38(02):796-806.DOI:10.13345/j.cjb.210166.
[11] 刘琦,毛雨丰,廖小平,罗家豪,马红武,姜文侠.麦角硫因生物合成研究的新进展[J].生物工程学报,2022,38(04):1408-1420.DOI:10.13345/j.cjb.210797.
[12] Liao, Wayne C., et al. “Kinetics of ergothioneine inhibition of mushroom tyrosinase.” Applied biochemistry and biotechnology 166 (2012): 259-267.
[13] Luisi, Grazia, et al. “Anti-oxidant and tyrosinase inhibitory in vitro activity of amino acids and small peptides: New hints for the multifaceted treatment of neurologic and metabolic disfunctions.” Antioxidants 8.1 (2018): 7.
[14] Chan, Chin-Feng, et al. “Inhibitory effects of novel synthetic methimazole derivatives on mushroom tyrosinase and melanogenesis.” Bioorganic & Medicinal Chemistry 22.9 (2014): 2809-2815.